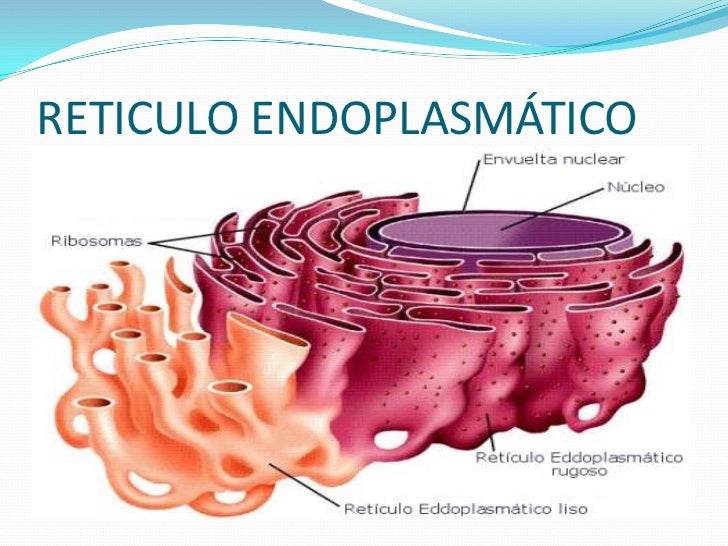
EL RETÍCULO ENDOPLASMÁTICO
Es una estructura membranosa multiplegada
dentro de las células eucariotas, que desempeña un papel importante en la
síntesis de las moléculas complejas requeridas por la célula y el organismo
como un todo. 
El retículo endoplásmico es un orgánulo
distribuido por todo el citoplasma de la célula eucariota. Forma parte del
sistema endomembranoso.
Existen
dos tipos de retículo endoplásmico que llevan a cabo funciones diferentes, el
rugoso y el liso.
El rugoso se encarga de la síntesis y el
plegamiento correcto de las proteínas mientras que el liso lleva a cabo la
síntesis de lípidos, almacenamiento de calcio y detoxificación de drogas
A menudo, las membranas de estas estructuras se
alinean con los ribosomas en sus superficies exteriores, dándoles una
apariencia áspera. Esta parte se llama retículo endoplásmico rugoso, en
contraste con el retículo endoplásmico liso, donde no hay ribosomas adjuntas.
Los ribosomas del retículo endoplasmático
rugoso fabrican proteínas que entran en los canales del retículo
endoplasmático, y los trasladan a lugares donde pueden crear bolsas. Estas
bolsas se pueden dividir como vesículas para transportar su carga de
proteínas al complejo de Golgi para su distribución.
Las proteínas producidas en las células
secretoras, son ejemplos de síntesis de proteínas por el retículo endoplásmico
rugoso.
Estas incluyen las enzimas digestivas
producidas en el estómago, y las hormonas de proteínas como la insulina
producida en el páncreas. Los sistemas de órganos que producen muchas
proteínas, tienen células con una gran cantidad de retículo endoplasmático
rugoso.
El retículo endoplásmico liso desempeña un
papel importante en la síntesis de los lípidos, por medio de enzimas
incrustadas en estas membranas lisas. Produce los fosfolípidos y
el colesterol utilizados en la formación de la membrana, y junto con las
proteínas de membrana producidas por el RE rugoso,
puede sintetizar más de
membranas por sí misma, para el complejo de Golgi, la membrana celular, los lisosomas,
y otros.
En las células del hígado el RE liso contiene
enzimas para la desintoxicación de drogas nocivas y de subproductos
metabólicos. En los órganos reproductivos, el ER liso en las células, produce
las hormonas esteroides testosterona, y estrógeno.
TIPOS
Es un orgánulo formado por una serie de túbulos,
sacos y vesículas rodeados de membrana e interconectados entre sí. Se pueden
distinguir dos tipos: el rugoso y el liso. En las células musculares se
encuentra un tercer tipo derivado del retículo endoplásmico liso conocido como
retículo sarcoplásmico.

RETÍCULO ENDOPLÁSMATICO RUGOSO
Presenta en su superficie ribosomas que se
encuentran sintetizando proteínas cuyo destino puede ser la membrana
plasmática, el exterior de la célula o los lisosomas y endosomas.
En el retículo rugoso
las proteínas que están siendo sintetizadas por los ribosomas se
pliegan y sufren también algunas modificaciones post-traduccionales como la
N-glicosilación sobre residuos de asparragina.
El plegamiento de las proteínas recién
sintetizadas es dirigido por las chaperonas. Las proteínas que se pliegan de
forma inadecuada son degradadas en un proceso conocido como UPR (Unfolded
Protein Response) o respuesta a proteínas mal plegadas. Fallos en esta
respuesta pueden causar el acúmulo de proteínas anómalas en el interior del
retículo que puede producir el llamado “estrés del retículo endoplásmico”.
Este tipo de estrés se relaciona con la
patogenia de importantes enfermedades como la diabetes o la ateroesclerosis. La
respuesta a proteínas mal plegadas está también relacionada con los procesos de
autofagia en la que se produce la degradación de los propios componentes de la
célula por formación de autofagosomas a partir de membranas del retículo
endoplásmico.
Tras su paso por el retículo endoplásmico las
proteínas pasan mediante vesículas a la cara cis del aparato de Golgi de donde
seguirán hacia su localización definitiva. Las proteínas residentes en el
retículo endoplásmico son recuperadas del aparato de Golgi mediante vesículas
que proceden de la cara cis del mismo y regresan al retículo.
Debido a su implicación en la producción de
proteínas que posteriormente serán secretadas el retículo endoplásmico rugoso
es muy abundante en células secretoras, como las células principales del
estómago o las células plasmáticas productoras de anticuerpos.

RETÍCULO ENDOPLÁSMATICO LISO
No presenta ribosomas. Sus funciones
principales son la síntesis de lípidos de membrana, el almacenamiento de calcio
y la detoxificación de drogas. Debido a esta última función, el retículo
endoplásmico liso es muy abundante en hepatocitos y aumenta con la ingesta de
sustancias tóxicas como el alcohol.
En células musculares lisas y estriadas
encontramos una forma especializada de retículo endoplásmico liso conocida como
retículo sarcoplásmico. El retículo sarcoplásmico es un importante almacén del
calcio que se utiliza en el proceso de contracción muscular.
CARACTERÍSTICAS GENERALES
· El
retículo endoplásmico se encuentra dentro de la célula, rodeado de membrana, y
establece dos compartimentos. Uno externo y otro interno.
· Está
en relación con otros orgánulos, constantemente en funcionamiento y dura toda
la vida celular.
· Se
encuentra en regiones basófilas del citoplasma (con microscopía óptica)
· Con
el M.E.T., el retículo endoplásmico está formado por:
2-Túbulos aplanados formados por una membrana
que rodea la luz (lumen) del retículo.
3-A la luz se le llama lumen o espacio
cisternal del retículo.
· El
retículo liso es muy poco abundante excepto en las células destinadas a
sintetizar hormonas esteroideas y al metabolismo alto de lípidos.
1-Queda reducido a una pequeña zona del
retículo elemento transicional.
· El
elemento transicional se caracteriza porque tiene ribosomas sólo en una cara.
· La
cantidad de retículo endoplasmático presente varia, depende del estado
metabólico de la célula.
· El
fenobarmital o los barbitúricos en células de rata aumentan el volumen del
retículo endopolasmático liso en mucha cantidad espuesta contra la toxicidad.
Si se deja de administrar se destruye por autofagia.
· El
retículo rugoso y el liso se aíslan fácilmente porque el retículo rugoso pesa
más.
1-Proteínas en las que se adhieren ribosomas
(en el rugoso)
· Las
dos membranas son similares. Formadas por:
FUNCIONES
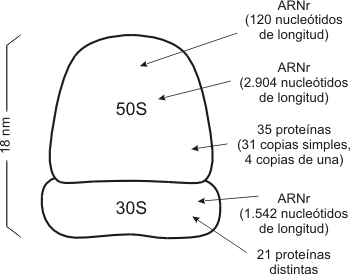
El ARN mensajero es el que lleva la información
para la síntesis de proteínas, es decir, determina el orden en que se unirán
los aminoácidos. Esta información está codificada en forma de tripletes, cada
tres bases constituyen un codón que determina un aminoácido.
Las reglas de correspondencia entre codones y
aminoácidos constituyen el código genético. La síntesis de aminoácidos tiene
lugar en los ribosomas del citoplasma.
Los aminoácidos son transportados por el ARN de
transferencia, específico para cada uno de ellos, y son llevados hasta el ARN
mensajero, dónde se aparean el codón de éste y el anticodón del ARN de
transferencia, por complementariedad de bases, y de ésta forma se sitúan en la
posición que les corresponde. Una vez finalizada la síntesis de una proteína,
el ARN mensajero queda libre y puede ser leído de nuevo.
De hecho, es muy frecuente que antes de que
finalice una proteína ya está comenzando otra, con lo cual, una misma molécula
de ARN mensajero, está siendo utilizada por varios ribosomas simultanéamente.
· Metabolismo
de lípidos: El retículo
endoplasmático liso, al no tener ribosomas le es imposible sintetizar proteínas
pero sí sintetiza lípidos de la membrana plasmática, colesterol y derivados de
éste como las ácidos biliares o las hormonas esteroideas.
· Detoxificación: Es un proceso que se lleva a cabo
principalmente en las células del hígado y que consiste en la inactivación de
productos tóxicos como drogas, medicamentos o los propios productos del
metabolismo celular, por ser liposolubles (hepatocitos).
· Glucosilación: Son reacciones de transferencia de un
oligosacárido a las proteínas sintetizadas. Se realiza en la membrana del
retículo endoplasmático. De este modo, la proteína sintetizada se transforma en
una proteína periférica externa del glucocálix en la reproduccion de lisosomas.